Alloreactivity is likely a term you have never heard of, but you are probably already somewhat familiar with the concept. If you have ever taken a biology class or donated blood then you may be aware that there are four different human blood types: A, B, AB, and O. Blood type is determined by the presence (or absence) of blood antigens on the surface of red blood cells. Antigen is just a fancy word for a protein that is recognized by the immune system, and in this case those proteins are the A and B proteins. You may express one blood antigen (type A or type B), both blood antigens (type AB), or neither of them (type O). The immune system recognizes these blood antigens on the surface of red blood cells and, if they are deemed “foreign” or unfamiliar, will mount an immune response against them. So, for safe transfusion of blood, certain blood types may only be given to people with a compatible blood type, although this does not necessarily mean that they must receive an identical blood type.
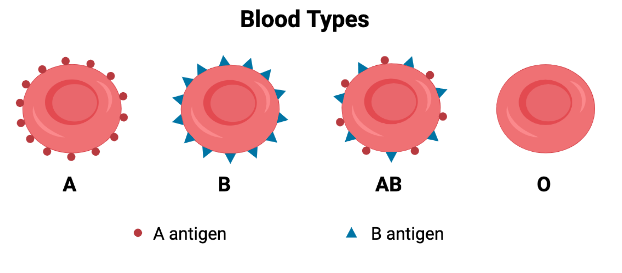
Type O blood, for instance, is considered the “universal donor,” meaning it can be safely given to anyone regardless of their blood type. This is because red blood cells from a type O individual do not have any blood antigens on their surface (no A protein and no B protein). Since type O blood lacks blood antigens, there is nothing to be recognized as foreign by the immune system, making it safe for anyone to receive. Alternatively, a person with type AB blood is considered a “universal recipient,” meaning they can accept blood of any type because their immune system is already familiar with both A and B antigens. As such, infusion of A, B, AB, or O blood in an individual with type AB blood will not cause any sort of adverse reaction. If you give incompatible blood however (e.g. type A blood given to a person with type B blood), that person’s immune system is only accustomed to seeing B antigen and will sense the A antigen as being a threat. This will cause the immune system to induce a severe immune reaction against all cells expressing A antigen, killing all the donor blood cells and simultaneously inducing an intense, and sometimes fatal inflammatory response.

Blood-typing is incredibly important to ensuring the safe transfusion of donor blood and is conceptually akin to alloreactivity. The word alloreactivity is derived from the Greek word “allo-“, meaning “other”, and refers to the ability of one’s immune system to recognize (and kill) cells that belong to someone else. Most of the time this is a non-issue, unless you require an organ transplant. When an organ in your body fails, sometimes the best course of action is to replace the failing organ with a new, functioning organ from a donor. Similar to blood-typing, in order for organ transplantation to be done safely it requires that the organ donor have a tissue type that is compatible with that of the organ recipient. This tissue type compatibility is known as histocompatibility (“histo-“ means tissue). Much like blood-typing, if you transplant an organ that is not compatible, the recipient’s immune system will recognize it as being foreign and kill it. This phenomenon is known as graft rejection and is one way that alloreactivity is manifested clinically.
Antigens on a Silver Platter (MHC)
While it is helpful to draw upon your knowledge of blood types to understand alloreactivity and histocompatibility, there are important distinctions to be made. While blood antigens (A and B proteins) may be present or not, tissue antigens are always present on all cells, tissues, and organs. These tissue antigens are immune proteins known as major histocompatibility complex (MHC) molecules. MHC molecules are incredibly important proteins for your immune health because they provide a platform for presenting materials to the immune system. Say you are infected with bacteria XYZ. Immune cells known as antigen-presenting cells (APCs) will capture the bacteria and break down XYZ proteins (the antigen) into smaller, bite-sized pieces of protein (i.e. peptides).
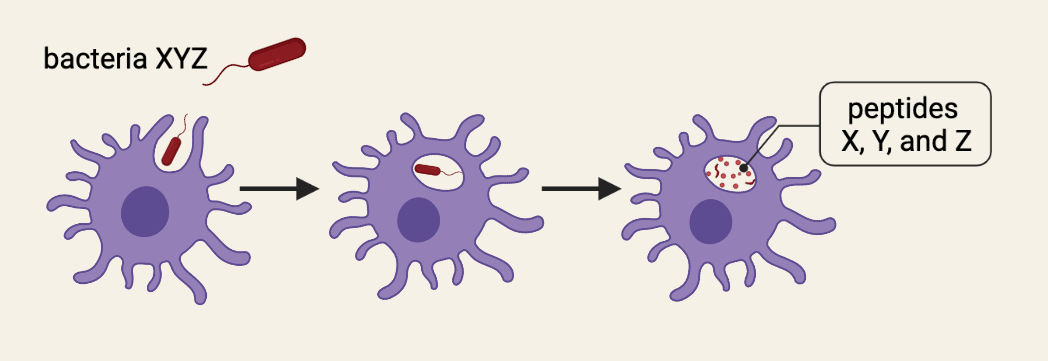
These X, Y, and Z peptides must then be presented to the T-cells of your immune system so that they can fight the infection. The problem is that your body contains millions of T-cells and, through a process known as V(D)J recombination, each T-cell has a unique T-cell receptor (TCR) that will only be activated by a very specific peptide. So, when APCs present X, Y, and Z peptides to T-cells, they run around from T-cell to T-cell looking for their one-in-a-million TCR (like Prince Charming looking for Cinderella using just a glass slipper). In order to make this process as efficient as possible, APCs “load” these peptides onto MHC molecules for presentation to T-cells. The MHC molecules essentially function as a silver platter for the T-cells to quickly check whether or not the peptide (“glass slipper”) fits.
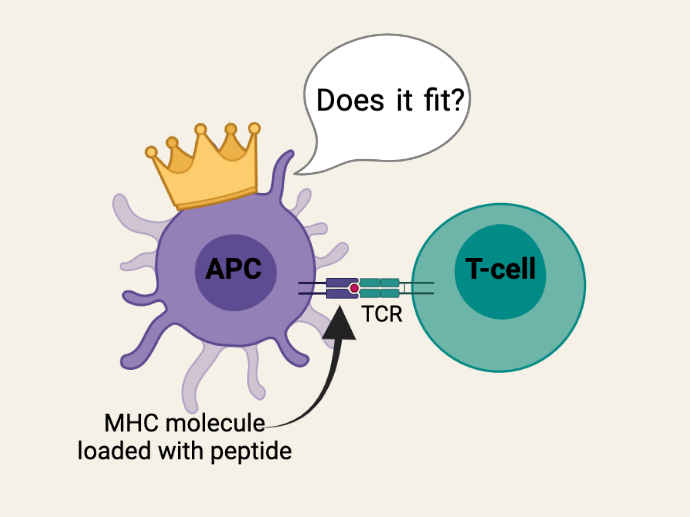
Because this is such an important aspect of your immune system’s ability to keep your body healthy, and an infection can happen anywhere in your body, MHC molecules are expressed by every single tissue and organ in your body. Unlike blood, there is no ‘universal donor’ or ‘universal recipient’ because there are no tissues or organs in the human body that do not express MHC molecules. This makes the task of finding a compatible organ donor much more challenging than finding compatible donor blood. Tissue-typing is required to find a donor who has MHC molecules that most closely match that of the transplant recipient; a greater disparity in MHC matching results in a greater risk of organ rejection (AKA graft rejection).
Allorecognition: What is it good for?
Alloreactivity initially seems intuitive–-of course our immune system knows when you introduce cells or tissue that belong to someone else. However, the evolutionary and molecular basis for alloreactivity has long been a subject of interest and debate among immunologists. Why? Because transplantation is a completely artificial biological process resulting from modern medicine. Organ transplantation and our understanding that tissue-typing is necessary to avoid graft rejection has only come to exist in the last ~200 years. But our immune system evolved over the course of millions of years, well before transplantation ever existed. So, why do we have this evolutionary mechanism for recognizing someone else’s cells? How does this benefit us?
One perplexing aspect of alloreactivity is the exceptional amount of genetic diversity of human MHC molecules (i.e. human leukocyte antigen; HLA). There are many different HLA types in humans and any single person can express up to 12 different versions. In other words, rather than just A and B antigens with blood-typing, HLA matching must consider A through L antigens for donor compatibility. This genetic diversity in MHC molecules is beneficial to human health because it allows a wider array of antigenic peptides to be presented. Having multiple HLA types in an individual allows Prince Charming to look for Cinderella (that one-in-a-million T-cell receptor) using a silver platter with her glass slipper (peptide loaded onto MHC), another platter with her glove, another platter with her sock, and so on. This increases the likelihood of finding a TCR that can strongly recognize (and kill) whatever pathogen is present, ultimately resulting in better immune responses.
But if there are millions of unique TCRs in a single person, and many different types of MHC molecules across the human population, then how are T-cells always able to recognize the MHC of another person? The first theory to explain alloreactivity was put forth by Niels Jerne in 1971 and posited that TCRs have segments that remain unchanged to ensure binding to MHC molecules. Jerne’s theory was eventually supported by the finding that the flank regions of the TCR (complementarity determining region (CDR)1 and CDR2) are germline-encoded, meaning they do not change during the process of making those millions of unique TCRs. These conserved regions ensure that every TCR can bind to the surface of any MHC molecule just enough to test out whether it is a perfect fit. The variable part of the TCR, CDR3, is unique to each T-cell and directly binds to the peptide embedded within the MHC molecules. The strength of CDR3 binding ultimately determines whether or not that T-cell crosses the threshold to become fully activated.
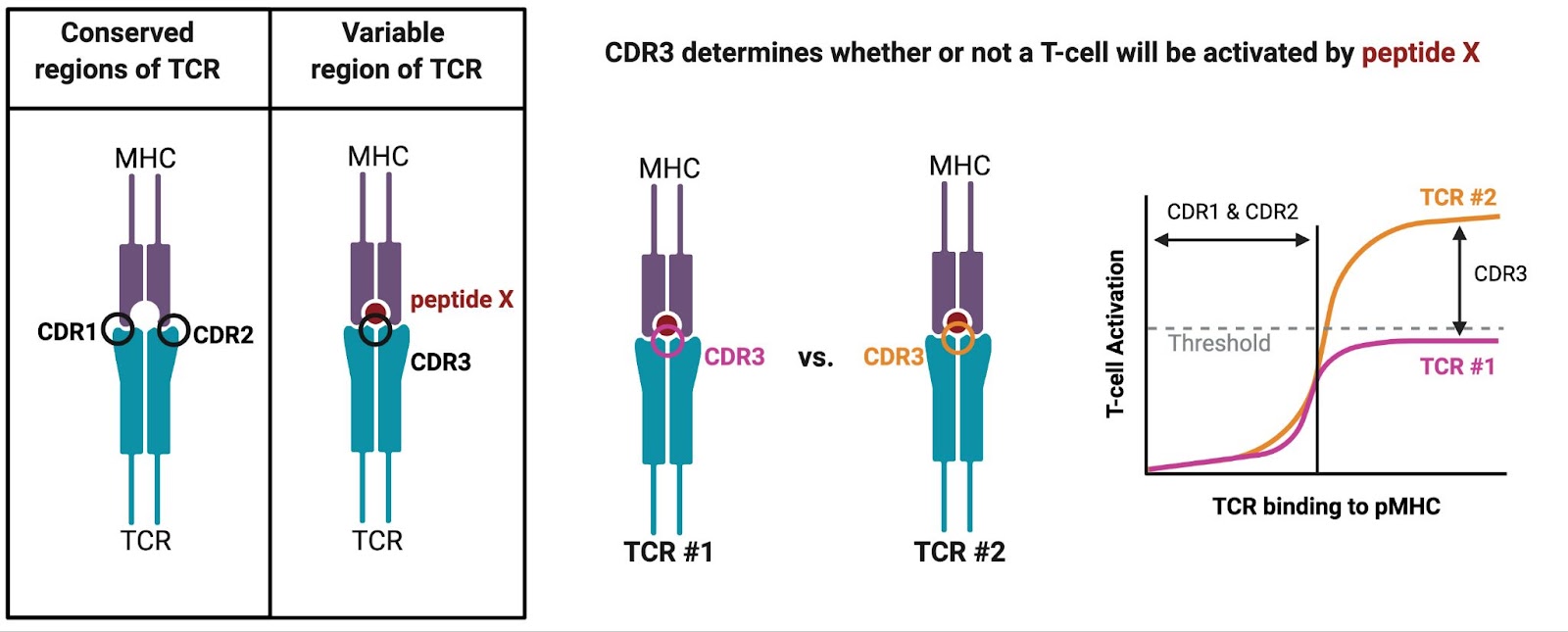
So, while MHC molecules and TCRs are both very diverse molecules, conserved structural pieces ensure that we have a functioning immune system with T-cells capable of efficiently recognizing antigens presented by MHC molecules. Germline-encoded MHC-binding may be deemed beneficial for immune health generally speaking, however it becomes problematic should one require an organ transplant, and especially if one requires a bone marrow transplant.
Bone Marrow Transplantation and Graft-versus-Host Disease
All the cells circulating in your blood come from stem cells in your bone marrow and they are constantly replenished throughout your lifetime. Just like with any other organ in your body, your blood may become diseased and need to be replaced with a healthy donor organ. Bone marrow transplantation (BMT) is used to treat various diseases of the blood, such as leukemia or aplastic anemia. Alloreactivity is much more evident in BMT and requires a much different way of thinking about alloreactivity compared to solid organ transplant. This is because all of your immune cells come from the bone marrow, so with a BMT you are essentially replacing the entire immune system, but most importantly are the T-cells. After infusion of healthy, donor bone marrow, the donor T-cells bind to the recipient’s MHC molecules via CDR1 and CDR2 and can mount an immune response.
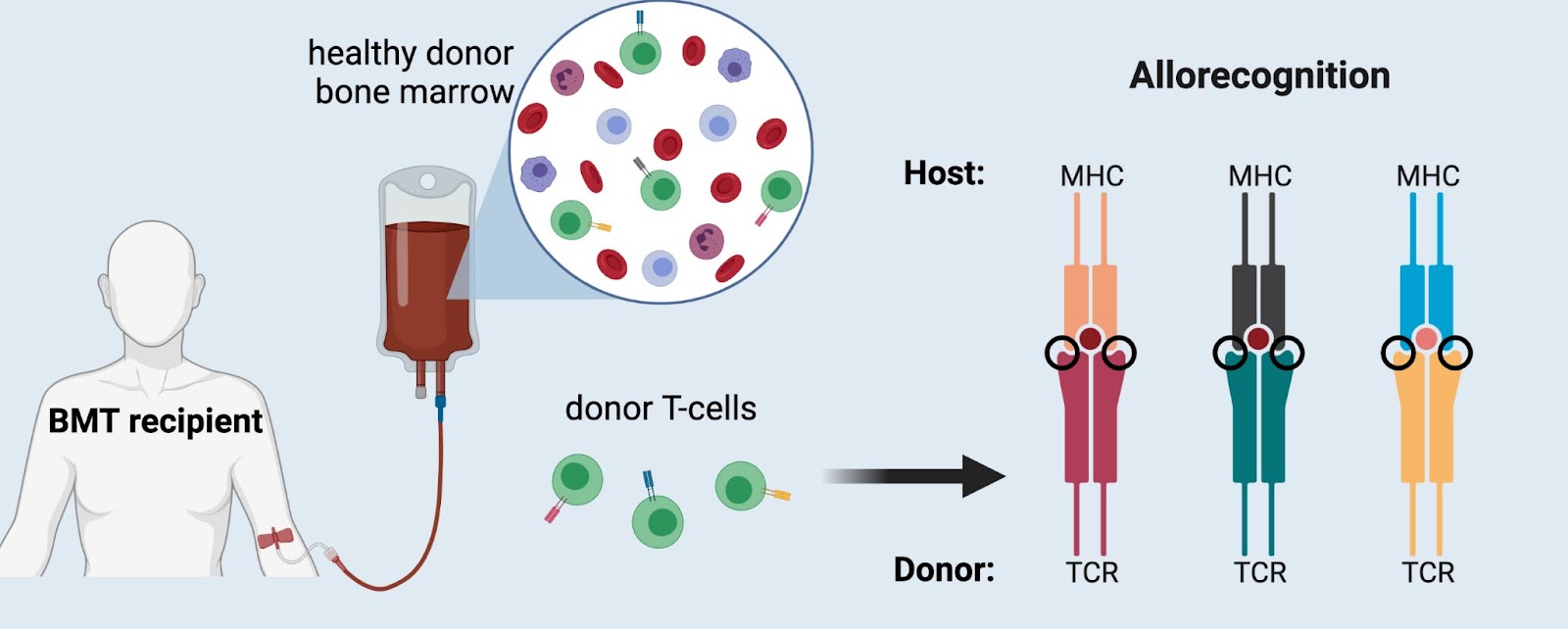
Importantly, however, this does not mean that every single donor T-cell will become activated. T-cell activation depends on how strongly a TCR binds to peptide-MHC; a very strong bond overcomes the activation threshold and allows that T-cell to divide and proliferate, whereas a weaker bond results in no immune response. We now understand that alloreactivity occurs when a TCR recognizes small differences in the protein structure (i.e. polymorphisms) in foreign MHC molecules. These small differences in MHC structure can sway the TCR to bind more strongly (think of Cinderella’s stepsisters forcing their feet into the glass slipper), resulting in inappropriate activation of the T-cell. Once a T-cell is activated it will proliferate and divide, creating many clones of itself which can then induce a robust, “alloreactive” immune response.
As opposed to solid organ transplant (where the alloreactive immune response can kill the donor organ and cause rejection), in BMT the alloreactive immune response is against the transplant recipient (i.e. the “host”). In this instance, there is not just a kidney’s worth of foreign MHC molecules, there is an entire body’s worth (skin, liver, intestines, lungs, etc.), providing an endless reservoir of foreign antigen. This culminates in graft-versus-host disease (GvHD), a severe immune response that attacks numerous tissues throughout the host body, most often the skin and intestines. Symptoms of GvHD include rashes, mouth ulcers, diarrhea, nausea, and anorexia. GvHD also prolongs the time to recovery for BMT recipients, meaning that they go for longer periods of time without any immune system whatsoever, contributing to opportunistic infections that can often be fatal. Something as simple as the common cold has the capacity to kill you when you don’t have an immune system.
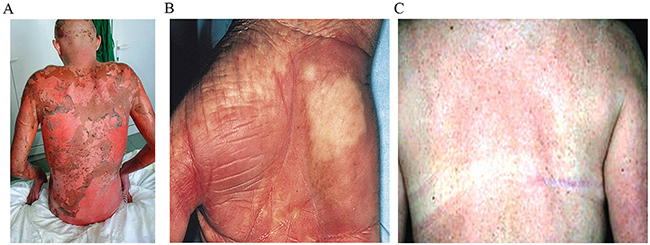
GvHD remains a significant obstacle in the transplant field and is the second leading cause of death in BMT recipients. Much like with solid organ transplantation, finding a compatible donor is among the largest determining factors for patient outcome, and the greater the disparity in HLA types between donor and recipient, the greater the risk of very severe, even lethal, GvHD. For this reason, the National Marrow Donor Program (NMDP) was created in 1986 to help BMT recipients find the best possible HLA-matched donor and reduce their risk of GvHD. This registry now maintains over 20 million adult volunteers and from it a global transplant network has emerged. Be The Match connects hundreds of transplant centers around the world and accepts cheek swabs from volunteers for HLA typing and registration into their database.
Tasmanian Devils & Contagious Cancer
While it seems as though alloreactivity does nothing more than make organ transplantation unbearably difficult, an interesting cautionary tale hails from Tasmania. Sadly, Tasmanian devils are facing extinction thanks to a highly contagious form of cancer known as devil facial tumor disease (DFTD). This transmissible cancer is spread from devil to devil via biting and is thought to result from a lack of allorecognition. The absence of allorecognition is primarily thought to be due to a lack of MHC diversity because of genetic bottlenecking in Tasmanian devil populations. Thus, when introduced with these tumor cells, they are not different enough from the Tasmanian devil’s own cells and are not recognized as foreign. This has allowed DFTD to spread rapidly and wreak havoc on the native populations of Tasmanian devils, which are now predicted to become extinct by the 2030s. Should humans ever become feral, face-biting animals, we can perhaps take solace in the fact that our genetic diversity and intact allorecognition mechanisms would prevent any sort of contagious cancer.
Image information:
Featured image:
“The Threatened Tasmanian Devil” by Pandora’s Perspective is licensed under CC BY-NC-SA 2.0
Figure 2:
“160113-F-YM354-011” by Armed Services Blood Program is marked with CC PDM 1.0
Figure 7:
http://dx.doi.org/10.18632/oncotarget.23276
All other figures created with BioRender.
Megan Stanley Molina is a Post-doctoral Scholar in the Department of Immunology at the University of Washington. Her research interests include innate control of adaptive immunity and cancer immunotherapy. You can find her on twitter @SciStan or instagram @meganst_nley.
