T cell receptors are required for the activation, regulation, and function of T cells. TCRs are generated by the random joining of gene segments in the TCR gene loci.
TCR assembly occurs through a process called V(D)J recombination, so named for the gene segments joined to make the β–chain of the TCR and the heavy chains of the B cell immunoglobulin receptor.
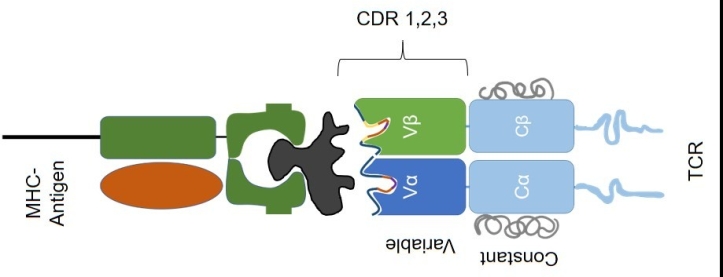
Figure 1: MHC-antigen complexes interact with the Variable-CDR regions of the T cell receptor.
The TCR interacts with antigen presented by MHC class I and Class II (Figure 1). The TCR is made up of Constant regions and Variable regions on both the alpha and beta chains (Figure 2, upper). Each Variable region is comprised of three complementarity-determining regions, or CDRs. Two of the three CDRs are mostly invariant and are derived from the germ line sequence (the genome) without alteration.
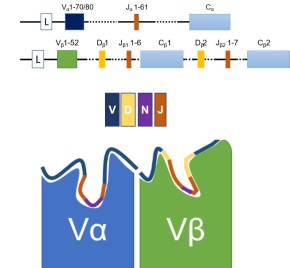
The antigen binding site and the site of maximum diversity is the CDR3 (Figure 2, lower). The CDR3 / antigen binding site interact with the peptides presented by MHC and are responsible for the specificity of T cells for their individual antigens.
The diversity of gene segments alone confers three million potential TCR combinations! Added to that are the diversity of random additions and excisions within the flanking regions of the gene segments.
In total, the estimated potential diversity of TCRs is 10^15-10^20.
Gene recombination is initiated in the beta-locus after thymic entry, when the T cell is double-negative for CD4 and CD8 markers. A complex molecular machine including proteins known as RAG, Artemis, and tDt (See the Nitty Gritty of TCR recombination in Immunology 201) cut, join, and prune the gene splice sites, creating a unique, part-genomic, part-random, new TCR gene segment.
Once a TCR β-chain has been successfully rearranged, the β-chain is paired to a pre-T-α chain. The pre-T chain allows the β-chain to correctly orient (extracelluar side out) and express in the plasma membrane with the TCR complex. This complex of pre-T, beta chain and signaling proteins is called the pre-TCR.
Expression of the pre-TCR initiates gene suppression of the beta-locus, rearrangement of the alpha locus and the expression of both CD4 and CD8 on the cell surface. For alpha-beta T cells (those discussed here), CD4 indicates T helper type cells, and CD8 expression pushes them into the cytotoxic T lymphocyte lineage.
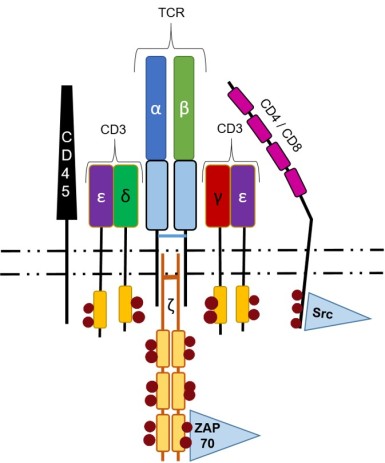
Multiple attempts can be made to generate a functional TCRα chain, and once accomplished, the T cells go on to undergo positive and negative selection before exiting the thymus. Approximately two percent of double-positive T cells will pass both selection steps and mature to single-positive, circulating T cells. The TCR complex (Figure 3) is a group of associated proteins required for signal transduction and signal propagation into the cell.

This site is so helpful. I’m self studying immunology and this site is honestly my savior!
LikeLike
We’re glad you’re learning something new! Thanks for the feedback.
LikeLike