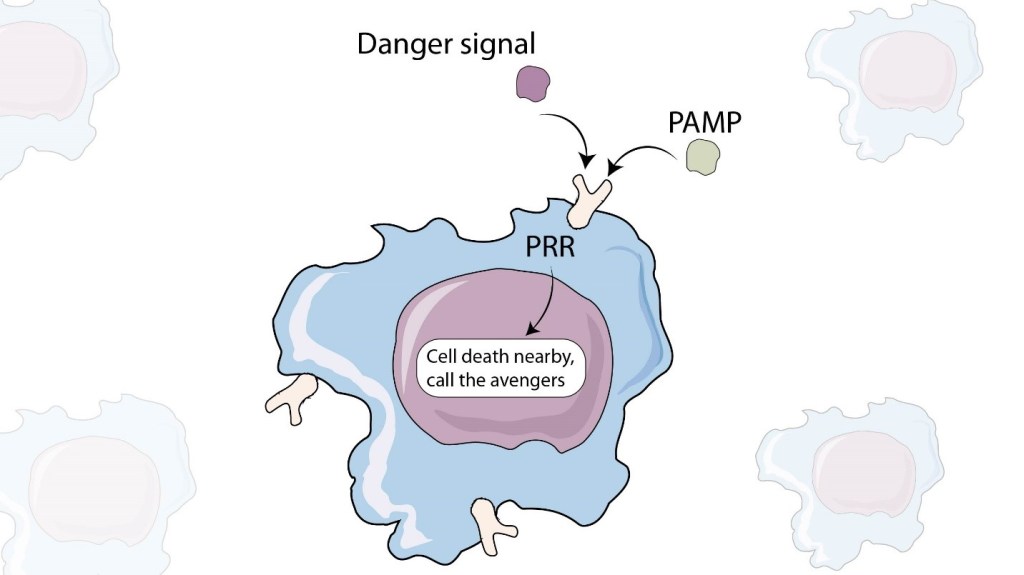
In 1994, it was postulated that our immune system does not respond to anything that is foreign, but specifically to substances that cause damage (Matzinger, 1994). To better understand this scenario, imagine that you see a stranger walking in front of your apartment complex, does that alert you? No. Now, imagine the stranger walking in front of your apartment complex has a knife that appears to have blood on it. This image suddenly puts you into an alert state and your “fight or flight” sense becomes heightened. Immune cells are also placed on high alert when they sense agents that cause cellular damage. This partly explains the reason that some of the most popular allergens are proteases, a class of enzymes that cleave proteins present on cell surfaces and which can look like cellular damage and put the immune system into an activated state. The cellular components and debris generated through these proteases are sensed by the immune system as danger signals through a set of receptors called pattern recognition receptors (PRR). Pattern recognition receptors recognize patterns of danger, e.g., extracellular ATP. Since ATP is an energy source for the cell, it is something that the immune system expects to only see inside the cell, never outside, unless the cell has burst open due to damage. Another example is cytoplasmic or extracellular DNA. DNA is also normally inside the nucleus of a healthy cell so its presence outside could indicate a damaged cell.
A common cell type that senses danger signals for the immune system is the dendritic cell (DC), which goes around gathering antigens. Sensing danger signals is required for the dendritic cell to mature, a process in which the DCs stop acquiring more antigens and enter nearby secondary lymphoid organs, like the lymph nodes, to present the antigens to T cells. This is a crucial role for DCs: mature DCs activate a helper T cell that recognizes exactly what antigen the mature DC is presenting. Once this antigen is recognized by the helper T cell, one of two processes can occur. First, helper T cells can provide support to cytotoxic T cells (cellular assassins of the immune system). This type of immunity is called cellular immunity. Second, helper T cells can activate B cells that will produce antibodies against the pathogen. This type of immunity is called humoral immunity. These processes are vital for generating a fine-tuned response against pathogens, cancer cells, and other threats in the body.
Pathogens can also directly activate PRRs via conserved structural elements on the pathogens called pathogen associated molecular patterns (PAMPs). This is another way that the immune system can recognize danger and race into action. But what if we look at this process through the eyes of the pathogen? A respiratory pathogen that infects the lungs should only cause damage in the lungs and would activate immune cells to migrate to the lungs and clear the infection promptly. However, PAMPs derived from this respiratory pathogen could also trigger the PRRs in other locations away from the lungs (liver, kidneys etc.) because they are dispersed throughout the body via the blood stream. In this case, it would make it difficult for the immune system to specifically detect the location of infection and most importantly, the location where the real damage is occurring. In this way, pathogen can try to throw the immune system off their scent so that they can continue to reproduce for as long as possible.
The most well studied PRR is called toll-like receptor 4 (TLR4). TLR4 senses lipopolysaccharide (LPS), an essential component of the cell membrane in Gram negative bacteria, a class of bacteria which contains many pathogenic species. LPS triggers TLR4 on host cells and alarms the immune system. Interestingly, TLR4 is a sensor for a host-derived protein as well that acts as a danger signal to alarm the immune system of cellular death (Gallucci & Matzinger, 2001). In a world where PRRs only recognized host-derived danger signals and not PAMPs, only pathogen-infected damaged cells would release danger signals and provide an exact location to where inflammation has occurred, possibly allowing a localized immune response and quick clearance. However in the world where PAMPs are recognized by PRRs, even dead or non-infectious bacteria can alarm the immune system and muddy the waters, making it harder for the immune system to pinpoint the site of infection. It is no surprise that PRR activation is directly linked to several autoimmune diseases where chronic inflammation guides our T cells to get rid of our own tissues (Hasham et al., 2017). We are often told that the immune system is diverse and smart. What we are not necessarily told is that pathogens are sometimes smarter.
Jatin Sharma is a graduate student at the University of Florida. He studies the protein “Suppressor of cytokine signaling-1” in context of autoimmune diseases like SLE and Psoriasis. When he is not in the lab, he is either podcasting, streaming video games on Twitch, or playing with his dog.
